| ЭЭГ корреляты медитативного состояния. | 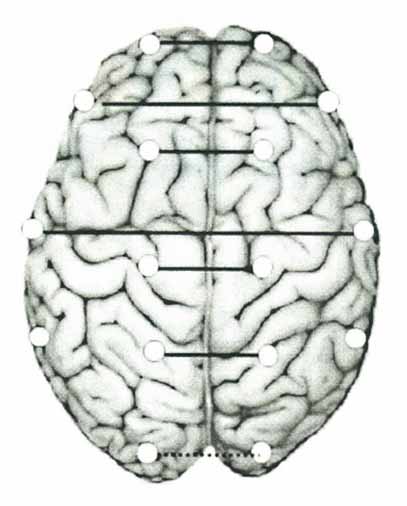 |
| назад... |
|
Пасынкова Н.Р. - кандидат биологических наук,
Метод отведения ЭЭГ При обычных измерениях ЭЭГ электроды помещаются на кожу черепа и мочки ушей. Электроды, помещаемые на кожу черепа, называются "активными электродами", а электроды, прикрепляемые к мочкам ушей, - референтными электродами. "Отведение" - это регистрация и запись разности потенциалов в парах электродов. Соединение электродов носит название "монтаж". Существует два типа монтажа: референтный и биполярный. Референтный монтаж предназначен для регистрации разности потенциалов между активными электродами, соответствующими каналам отведения ЭЭГ, и референтным электродом. Биполярный монтаж предназначен для регистрации разности потенциалов между любыми двумя активными электродами без использования референтного электрода. Последний метод эффективен при локализации очагов аномальной ЭЭГ. Метод локализации Локализация - это нахождение электрода, расположенного ближе всех остальных к району возникновения определенной активности ЭЭГ. Когда четкий образец ЭЭГ локализуется в определенном электроде, можно считать, что активность возникает в мозговой ткани вблизи электрода. Если, например, на каком-то электроде фиксируются аномальные изменения потенциала, как на графике 6, то источник этой аномальности легко локализовать. Локализация сводится : 1)K определению источника той или иной активности ЭЭГ (например, пиковые или медленные волны). 2)K определению области, в которой происходит возрастание/убывание амплитуды обычных феноменов ЭЭГ (альфа-волны или пиковые волны), или района, где активность ЭЭГ отсутствует. Мозговая электрическая активность, регистрируемая ЭЭГ электродами Поскольку всякий компонент мозга частично входит в кору головного мозга, состояние этих компонентов может быть зафиксировано на ЭЭГ кожи черепа, в основном записывающей активность коры головного мозга. Однако, ЭЭГ кожи черепа может и не показать никаких признаков аномальности, возникающей в глубине коры головного мозга или в мозговом стволе. Так что, ЭЭГ кожи черепа ограничена. Рисунок показывает соответствие между расположением электродов на коже черепа и находящимися под ними функциональными образованиями мозга. Характеристики типичных образцов ЭЭГ, записанных с некоторых основных электродов, могут быть вкратце выражены следующим образом: FР1 . FР2 (фронтополярные): лобные ассоциативные зоны и лимбическая система. По данным клинической неврологии это спокойная зона, где сложно обнаружить отклонения. Симптомы в психологическом плане: иногда возникает безрассудство влодедении. Альфа- волны слабы даже в ЭЭГ обычного типа, соответствующей состоянию спокойного бодрствования. В этой зоне во время некоторых сложных частичных и побочных приступов, сопровождаемых вращением головы и глазных яблок, фиксируются пароксизмы. С3, С4 (центральные): обнаруживают отклонения в моторной и соматической сенсорных областях, так как они расположены над центральной бороздой. Р3, Р4 (теменные): соответствуют лобным ассоциативным зонам. Этот район отвечает за разнообразные симптомы агнозии и апраксии. О1, О2 (затылочные): содержат зрительную зону и окружающие ее зрительные ассоциативные зоны, а также район, контролирующий движение глаз. Т3, Т4 (центральные височные): также входят ассоциативные зоны. Внутри расположены гиппокамп и миндалевидные ядра клеток. Когда в височных ассоциативных зонах или гиппокампе возникает эпилептогенный очаг, судорожные припадки, сопровождаемые такими трансформациями сознания, как иллюзии, галлюцинации, провалы в памяти,перемена эмоций. На вершине височной доли расположен сенсорный речевой центр Вернике. F7, F8 (Фронтальные височные): несмотря на свое расположение в лобной доле, эти электроды близки к глазному мозгу, который относится к лимбической системе. Они отражают деятельность фронтальной височной доли в большей степени, чем деятельность лобной доли. Фронтальная височная доля, содержащая в себе миндалевидные ядра клеток, - это очень важная часть. Это область, в которой наиболее часто возникают очаги сложных частичных приступов, в особенности очаги эпилептогенной автоматии. Несмотря на большие достижения в области нейронаук, Сознание остается одной из самых непознанных функций мозга. Последнее десятилетие XX века ЮНЕСКО предложила назвать десятилетием "Сознания". Пришло время с естественно научных физиологических позиций заглянуть в эту величайшую тайну Вселенной. Существующие научные подходы к изучению этой проблемы разнообразны. В этой связи нам представляется целесообразным изучение такого феномена или особого состояния сознания, когда в глубокой интроспекции разум познаёт самое себя. Подобный феномен в нейропсихологии называют инсайтом, или озарением, или самоосознанием. В обыденной жизни такие феномены редки, спонтанны, и в связи с этим трудны для изучения. Но оказалось возможным воспроизвести подобные состояния и зафиксировать их в экспериментальной ситуации, с помощью метода медитации по системе Сахаджа Йоги. Почему из всех возможных школ, практикующих медитацию, медитация по системе Сахаджа Йоги является наиболее перспективной для изучения подобного феномена? По ряду причин. I. Во время медитации по системе Сахаджа Йоги достигается состояние осознания без мысли, то есть без участия речевых функций, без внутреннего монолога и без зрительных образов. В нейронауках обычно сознание и речь или зрительный образ рассматривают как тесно сопряженные функции. Работая с моделью Сахаджа Йога медитации, мы имеем дело, так сказать, с чистым феноменом сознания, без участия зрительных и речевых функций. II. Кроме того, само понятие медитации не однородно. Разные течения предлагают различные подходы для достижения медитативного состояния. Но суть этих различий заключается в воздействии на разные сферы психической деятельности человека. В настоящее время общепринятой в науке является точка зрения, что психическая деятельность не исчерпывается лишь функцией сознания. Структура психического включает также сферу подсознания, которая связана с индивидуальным и коллективным прошлым, и сферу сверхсознания, связанную с будущим. В большинстве школ, практикующих медитацию, воздействие направлено для достижения состояния подсознания или сверхсознания. В современной европейской психологии довольно хорошо описаны способы воздействия или самовоздействия для погружения в трансовые состояния, связанные со сферой подсознания или сверхсознания. Такие состояния могут быть неконтролируемыми и приводить к болезненным изменениям. В психиатрии известны такие случаи. Сейчас многими современными психиатрами признается тот факт, что некоторые психические заболевания связаны с неконтролируемым погружением в сферу под- или сверх-сознания. В противовес этому, медитация по системе Сахаджа Йоги включает работу со сферой Сознания, связанной с настоящим "здесь" и "сейчас", и это состояние является полностью контролируемым. III. И последняя немаловажная причина интереса нейрофизиологов к Сахаджа Йога медитации - это то, что медитативное состояние может быть достигнуто и воспроизведено многими людьми, практикующими Сахаджа Йогу. Кроме того, практика эта не ведет к пагубным последствиям, а напротив, оказывает благотворное воздействие на организм человека, что показано во многих работах. Целью данного исследования было изучение электроэнцефалографических (ЭЭГ) особенностей, характерных для глубокого медитативного состояния. В эксперименте участвовали 10 сахаджа йогов, практикующих Сахаджа Йогу более 5 лет и восемь контрольных испытуемых - студентов университета и сотрудников НИИ, не практикующих йогу, которые находились в сходной экспериментальной ситуации, для выяснения влияния частичной сенсорной депривации на изменение ЭЭГ. Аппаратура и процедура. Эксперименты проводили в изолированной экранированной камере. В ходе эксперимента первоначально была сделана фоновая запись ЭЭГ в состоянии спокойного бодрствования с закрытыми глазами длительностью 200 сек., далее во время медитации, которая продолжалась около 30 мин., регистрировалась ЭЭГ в виде отдельных реализаций длительностью 200 сек. с перерывами между регистрациями около 60 сек. При выходе из медитативного состояния также была сделана фоновая запись. Во время записи ЭЭГ контрольные испытуемые находились в состоянии спокойного бодрствования с закрытыми глазами в течении 30 мин. ЭЭГ регистрировалась в начале эксперимента, в середине, то есть через 14 минут после начала эксперимента, и в конце, то есть спустя 27 минут от начала. Длительность каждой регистрации ЭЭГ составляла 180 секунд. ЭЭГ регистрировали от 16 отведений, расположенных в симметричных точках правого и левого полушарий по системе 10/20, от передне-фронтальных (Гр1-Гр2), фронтальных (ГЗ-Г4), центральных (СЗ-С4), париетальных (РЗ-Р4), окципитальных (О1-О2), фронто-темпоральных (Г7-Г8), темпоральных (ТЗ-Т4), и заднетемпоральных (Т5-Т6); с объединённым ушным референтным электродом. Анализ ЭЭГ осуществлялся с помощью компьютерной программы "Нейрокартограф-3.55". Для каждого отведения методом быстрого преобразования Фурье были получены характеристики мощности для диапазонов: дельта (0,5-4 Гц), тета (4-8), альфа (8-13), по которым методом интерполяции строились карты распределения спектров мощности. На основе индивидуальных карт строились групповые карты распределения спектров мощности в начале эксперимента в середине и в конце. Для изучения изменений спектров мощности ЭЭГ в разные фазы эксперимента проводили статистический анализ методом критерия Стьюдента. Когерентность анализировали между симметричными отведениями правого и левого полушарий мозга. Статистический анализ проводили по методу Манна-Уитни. Рис. 1 Спектр ЭЭГ Мощности: 1 - фоновая активность мозга в состоянии спокойного бодрствования; 2, 3, 4 - в состоянии медитации; 5 - ритмы мозга после выхода из медитативного состояния. На рисунке 1 представлены результаты картирования мозга по спектру мощности, полученные по пяти реализациям: первой - в состоянии спокойного бодрствования, трем реализациям в состоянии медитации, и пятой в состоянии спокойного бодрствования по выходе из медитации. Максимальные значения на рисунке представлены красным цветом, минимальные - синим (см. цветовую шкалу в нижней части рисунка). Результаты анализа мощности ЭЭГ показали, что во время медитации изменяется спектральная мощность во всех исследуемых диапазонах. В первом столбце карт спектральной мощности на рис.1 представлены данные фоновой активности мозга в состоянии спокойного бодрствования, из которых видно преобладание альфа ритма в спектре, с преобладанием его в правой затылочной области, что является среднестатистической нормой. С началом медитации ЭЭГ паттерн значительно изменяется (см. 2-й столбец карт): снижается мощность альфа ритма в затылочной области и увеличивается выраженность медленных ритмов - дельта и тета, преимущественно в центральных отделах. С углублением медитативного состояния (см. 3-й и 4-й столбцы) постепенно нарастает спектральная мощность медленных ритмов. К концу эксперимента при выходе из медитативного состояния мощность ЭЭГ приближается к исходному уровню (5 столбец). На основании данных о природе электрической активности мозга [а именно, в общих чертах: более быстрые волны (бета, альфа) генерируются в более поверхностных структурах мозга, более медленные (тета, дельта) в более глубоких, кроме того, известно, что кора принимает участие в модуляции всех корковых ритмов], можно сказать, что резкое снижение мощности альфа ритма в начале медитации отражает реакцию активации. По мере углубления медитативного состояния, наблюдается рост мощности тета и дельта ритмов. Учитывая локализацию в центральных областях, эти изменения можно интерпретировать как усиление активности подкорковых лимбических структур. Далее, в состоянии глубокой медитации максимальная выраженность тета и дельта ритмов и незначительный рост альфа указывают на то, что наряду с высокой активностью подкорковых структур, усиливается модулирующая функция коры мозга, что, вероятно, является нейрофизиологическим отражением того, что медитация в Сахаджа Йоге является контролируемым процессом. При этом, однако, основные функциональные изменения происходят в подкорковых структурах, то есть независимо от таких контролируемых корой функций, как представления, установки. Снижение мощности альфа ритма в затылочной области во время медитации, вероятно связаны со снижением функциональной активности зрительной затылочной коры, которая по данным Изнак А. Ф. (1988) является независимым от таламического источником генерации альфа ритма. Рис. 2 Изменение когерентности ЭЭГ во время медитации. Сплошные линии - увеличение когерентности; пунктирная линия - снижение когерентности. На рисунке 2 представлены результаты анализа когерентности ЭЭГ во время глубокой медитации, по сравнению с фоновой записью. Сплошные линии на рисунке означают увеличение когерентности между двумя отведениями, пунктирные - снижение. Было обнаружено, что во время глубокой медитации увеличивается когерентность во фронтальных и центральных отделах и снижается в затылочных. Согласно современным представлениям, функция когерентности отражает степень сонастроенности мозга в реализации психофизиологического процесса. Более высокие значения когерентности между двумя сигналами, записанными от двух симметричных областей мозга означают более высокую степень интеграции или большее функциональное взаимодействие между этими корковыми областями. Рост когерентности в состоянии глубокой медитации в передних и центральных отделах мозга, что свидетельствует о более высоком функциональном взаимодействии полушарий мозга, причем его передних отделов. Известно, что передняя кора принимает участие в сложных интегративных процессах. Известно также, что фронтальные отделы в тесной связи с лимбическими структурами, принимают участие в эмоциональной регуляции. Кроме того, рост межполушарной когерентности ЭЭГ во фронтальных и центральных отделах отражает увеличение функциональной активности подкорковых структур мозга и большую степень кооперативного взаимодействия коры и подкорковых структур. Снижение когерентности в затылочной области, по-видимому, указывает на снижение функциональной активности зрительной коры. В группе контрольных испытуемых достоверных различий обнаружено не было. Таким образом, на основании проведенного исследования можно сделать следующие выводы. Во время глубокой медитации по системе Сахаджа Йоги: усиливается функциональная активность подкорковых лимбических структур, растет степень кооперативного взаимодействия коры мозга с подкорковыми структурами, увеличивается функциональное взаимодействие полушарий мозга в передних и центральных отделах, снижается уровень функциональной активности зрительной коры. Научная новизна работы. Впервые в системном психофизиологическом исследовании с использованием ЭЭГ высокого пространственного разрешения обнаружены объективные корреляты изменённого состояния сознания в условиях Сахаджа йога медитации. Показано, что изменённое состояние сознания в условиях медитации характеризуется усилением локальной тета-активности избирательно в передне-медиальных отделах коры головного мозга и ростом длиннодистантной тета коннективности между префронтальной и задней ассоциативной корой. Обнаруженные объективные изменения значимо коррелируют с возникающими в состоянии медитации положительными эмоциональными переживаниями. Впервые установлено, что восприятие эмоциональных стимулов медитаторами характеризуется более "экономным" использованием активационных ресурсов. Данный феномен отражается в более низких значениях субъективной оценки эмоционального содержания стимула и в менее выраженных эффектах вызванной синхронизации мощности ЭЭГ в дельта-, тета-1-, тета-2- и гамма-диапазонах. Впервые показано, что на субъективном уровне медитаторы демонстрируют меньшую интенсивность переживания вызванных дискретных эмоций. На нейрофизиологическом уровне, особенности реализации эмоций отражаются в изменениях реактивности в тета-2-, бета-1-, бета-3- и гамма-диапазонах ЭЭГ. Впервые в модели лабораторного психоэмоционального стресса обнаружено, что защитные эффекты медитации проявляются в менее выраженных эффектах симпатической активации в ответ на стрессовый вызов и связаны с изменениями активности ЭЭГ в бета-1-диапазоне. Представленные в работе результаты включены в лекционные курсы по психофизиологии Новосибирского государственного университета и Новосибирского государственного педагогического университета. Полученные психофизиологические данные учитываются при построении тактики лечения больных ряда психосоматических заболеваний в муниципальной клинической поликлинике № 21 г. Новосибирска. Испытуемые. В исследовании приняли участие 93 здоровых добровольца (правши) в возрасте от 20 до 40 лет. Из них 43 (22 мужчины, 21 женщины) составляли группу контрольных испытуемых, не практикующих медитацию либо другие техники психической саморегуляции, а 50 (24 мужчины, 26 женщин) - экспериментальную группу медитаторов, регулярно практикующих Сахаджа йога медитацию. Перед началом исследования у испытуемых обеих групп оценивали уровни личностной тревожности (STAI-t) (Ханин 1989, Spielberger 1983), алекситимии (TAS-20) (Bagby 1994), депрессии (BDI) (Beck et al., 1961), а также экстраверсии/интроверсии, нейротизма и психотизма (EPQ) (Hanin et al., 1991). Экспериментальные модели. 1) Состояние физиологического покоя. Испытуемые располагались в удобном кресле в затемнённой комнате. Состояние физиологического покоя регистрировали при открытых и закрытых глазах (ОГ и ЗГ соответственно) в следующей последовательности: ОГ (90 с.), ЗГ (90 с.), ОГ (90 с.), ЗГ (180 с.). 2) Модель изменённого состояния сознания. Ключевыми элементами техники Сахаджа Йога медитации является погружение в состояние, связанное с временной "остановкой мыслей", т.е. прекращения когнитивной активности и возникновением интенсивных положительных эмоциональных переживаний, категоризуемых как "счастье" или "блаженство". В процедуре медитации было выделено три последовательные стадии, во время которых регистрировались ЭЭГ: 1) вхождение в медитацию (5 мин.); 2) глубокая медитация (15-20 мин.); 3) выход из состояния медитации (5 мин.). После эксперимента каждый испытуемый при помощи 9-балльных шкал оценивал интенсивность положительных эмоциональных переживаний и мыслительной активности во время медитации. 3) Модель восприятия эмоциональной информации. В работе использованы зрительные стимулы Международной аффективной системы изображений (IAPS, Center for the Study of Emotion and Attention, CSEA-NIMH, 1999), которые были разделены на 3 категории: нейтральные, эмоционально положительные и отрицательные (по 30 стимулов на каждую категорию). Экспериментальные стимулы предъявляли на мониторе стимулирующего компьютера (время экспозиции - 500 мс.). Для каждого стимула общее время регистрации ЭЭГ составляло 8500 мс. (6500 мс. до и 2000 мс. после начала предъявления стимула). По окончании предъявления каждого стимула испытуемый оценивал эмоциональную категорию сигнала (приятный, неприятный, нет эмоций) и нажимал на соответствующую кнопку обратной связи. Стимулы предъявлялись в псевдослучайном порядке. Эксперимент состоял из 90 предъявлений, разделённых на два блока по 45 предъявлений в каждом. В конце эксперимента испытуемые повторно просматривали все изображения и оценивали своё эмоциональное впечатление по 9-бальным шкалам: 1) "приятно - неприятно"; 2) "спокоен(а) - вобужден(а)". 4) Модель переживания вызванных дискретных эмоций. Для динамической эмоциогенной стимуляции использовались калиброванные по интенсивности и знаку эмоционального контекста аффективные видео клипы, сбалансированные по элементам сценария. Все клипы представляли собой фрагменты цветных, звуковых видеофильмов длинной 3 - 4,5 мин. Два эмоционально нейтральных видеофрагмента использовались в качестве референтного условия. Тестовые аффективные видео клипы индуцировали различные эмоциональные состояния: релаксации, радости, гнева, отвращения, грусти, психоэмоционального стресса. Одновременно с предъявлением видео клипов на экране монитора регистрировалась ЭЭГ. Для каждого тестового эмоционального состояния отбирали по 3 фрагмента ЭЭГ (8192 мс.) с учётом реакций электрокожного потенциала (ЭКП). По окончании предъявления каждого клипа испытуемый оценивал вызванные дискретные эмоции по 9-бальным шкалам. РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ 1) Данные психометрии и спектры мощности ЭЭГ в состоянии физиологического покоя. А) По данным психометрии, медитаторы по сравнению с контролем характеризовались более низкими значениями экстраверсии (I: 11,77 vs. 14,12, p<0,029), нейротизма (II: 3,070 vs. 7,820, p<0,001), психотизма (III: 3,230 vs. 4,780, p<0,014), личностной тревожности (IV: 35,10 vs. 41,72, p<0,001), депрессивности (V: 2,230 vs. 8,000, p<0,001), а также меньшим количеством баллов в субшкалах трудности в идентификации чувств (VI: 12,85 vs. 16,65, p<0,002) и выражения эмоций (VII: 10,59 vs. 12,96, p<0,032) по данным опросника на алекситимию (см. рис. 1) Рис. 1. Данные психометрии. Б) Показатели активности вегетативной нервной системы. Данные корреляционной ритмографии сердечного ритма показывают, что отношение длинной оси скатерограммы к короткой (L/W) у медитаторов значимо больше (I: 2,964 vs. 2,337, p<0,001). Анализ нелинейных оценок сердечного ритма обнаруживает достоверно более высокие уровни фрактальной размерности у медитаторов (II: 5,166 vs. 4,615, p<0,003). В то же время амплитуда спонтанных реакций кожного потенциала (III: 0,615 vs. 0,979, p<0,036) и частота дыхания (IV: 11,97 vs. 14,20, p<0,014) у медитаторов оказалась ниже, чем у контрольных испытуемых (см. Рис. 2) Рис. 2. Показатели активности вегетативной нервной системы. В) Спектральная мощность ЭЭГ. Только в состоянии покоя с закрытыми глазами группа медитаторов по сравнению с контролем характеризуется более высокими значениями мощности в тета-1- (F(1,55)=5,549, p<0,002), тета-2- (F(1,55)=5,063, p<0,028) и альфа-1- (F(1,55)=5,495, p<0,023) диапазонах. При этом, в тета-1- и тета-2- диапазонах преобладание мощности наблюдается в передне-центральных, а в альфа-1-диапазоне - во всех корковых областях. В то же время в гамма-диапазоне как при закрытых, так и открытых глазах у медитаторов наблюдаются более низкие значения мощности во всех областях коры (F(1,55)=4,578, p<0,037). Таким образом, по данным психометрии медитаторы по сравнению с контрольными испытуемыми, обладая большей интровертированностью, характеризуются более высокой психоэмоциональной стабильностью и лучшими способностями к идентификации и выражению эмоций. На вегетативном уровне по данным анализа частоты сердечных сокращений медитаторы характеризуются меньшими значениями симпатической активации и более выраженной вариативностью сердечного ритма. Более низкая амплитуда спонтанных реакций ЭКП также указывает на меньшую тоническую активность симпатической нервной системы. По данным ЭЭГ медитаторы демонстрируют более высокие значения тета-1- и тета-2-мощности в передних областях коры. В альфа-дипазоне медитаторы характеризуются более высокими, а в гамма-диапазоне более низкими значениями мощности во всех областях коры. 2) Спектры мощности, когерентность и фрактальная размерность ЭЭГ в состоянии медитации. А) Субъективный отчёт. Согласно данным субъективных оценок медитаторы достигали значимо большей, чем контроль, интенсивности переживания состояния "блаженства" (5,54 vs. 3,56, p<0,014) и более низкого уровня мыслительной активности (1,19 vs. 2,28, p<0,025). Б) Спектральная мощность ЭЭГ. При сравнении состояния покоя и медитации для симметричных корковых зон левого и правого полушария было обнаружено значимое взаимодействие факторов ГРхЭУ в тета-2- (F(1,25)=5,299, p<0,030) и альфа-1- (F(1,25)=5,923, p<0,022) диапазонах, а так же ГРхЭУхЛОК в альфа-2-диапазоне (F(5,125)=5,914, p<0,010). У медитаторов наблюдался рост мощности в тета-2- и альфа-1- диапазонах преимущественно в передних отделах коры, а в альфа-2-диапазоне - в задних отделах коры. Контрольные испытуемые демонстрировали снижение мощности в альфа-1- и в альфа-2- диапазонах в задних отделах коры. При анализе значений спектральной мощности в электродных кластерах средней линии также было обнаружено взаимодействие ГРхЭУ в тета-2- (F(1,25)=5,266, p<0,030), альфа-1- (F(1,25)=4,526, p<0,043) диапазонах и ГРхЭУхЛОК в тета-1- (F(5,125)=4,112, p<0,029), альфа-2- (F(5,125)=3,759, p<0,030) диапазонах. У медитаторов наблюдался рост тета-1-, тета-2- и альфа-1-мощности в передне-медиальных зонах, а альфа-2-мощности в затылочно-медиальной зоне. В результате корреляционных анализов были обнаружены значимые связи между изменениями спектральной мощности в тета-1-, тета-2-, альфа-1- и бета-1-диапазонах и оценками субъективных характеристик медитации. Интенсивность положительных эмоциональных переживаний в условиях медитации положительно коррелирует с ростом тета-1-мощности в передних и центральных корковых зонах (значимые корреляции в диапазоне от r=+0,38 до r=+0,41). В свою очередь, интенсивность мыслительной активности отрицательно коррелирует с изменениями тета-2-мощности (значимые корреляции от r=-0,38 до r=-0,52) по всему корковому плану и альфа-1- (от r=-0,38 до r=-0,52), бета-1-мощностью (от r=-0,45 до r=-0,49) в передних отведениях. В) Когерентность ЭЭГ. Эффекты медитации были обнаружены только в тета-1-диапазоне (4-6 Гц). В условиях медитации группа медитаторов характеризовалась усилением коротко- и длиннодистантных когерентных связей преимущественно в передне-заднем направлении с выраженным "центром тяжести" левой префронтальной области. Г) Фрактальная размерность ЭЭГ. В условии медитации медитаторы демонстрируют фокусированное снижение оценок DCx в передних отведениях средней линии, что подтверждается значимым взаимодействием ГРхЭУ (F(1,25)=4,982, p<0,036) и результатами последующего внутригруппового анализа для группы медитаторов (F(5,75)=5,400, p<0,012). У медитаторов оценки DCx и значения мощности средней линии отрицательно коррелировали между собой в тета-2- (от r=-0,52 до r=-0,57) и альфа-1- (от r=-0,56 до r=-0,66) диапазонах. Таким образом, состояние медитации характеризуется усилением тета-1-, тета-2- и альфа-1-мощности в передних областях коры, наряду со снижением оценок фрактальной размерности, а в задних областях коры - ростом альфа-2-мощности. Рост тета-1-мощности положительно коррелирует с выраженностью положительных эмоциональных переживаний, а изменения тета-2-, альфа-1- и бета-1-мощности отрицательно коррелирует с интенсивностью мыслительной активности. Кроме того, состояние медитации сопровождается ростом длиннодистантной тета-1-когерентности между префронтальной и задней ассоциативной корой с формированием "центра тяжести" в левой префронтальной области. 3) Вызванная синхронизация и десинхронизация в процессе опознания эмоциональности сигнала. А) Субъективный отчёт В оценке знака (F(2,80)=18,479, p<0,001) и активационного содержания (F(2,80)=18,479, p<0,001) стимулов обнаруживаются отчётливые межгрупповые различия - медитаторы по сравнению с контрольными испытуемыми оценивают положительные и отрицательные стимулы как менее эмоциогенные. Б) Данные вызванной синхронизации и десинхронизации. Вне зависимости от категории стимула, медитаторы характеризуются меньшими значениями ВС в дельта- (F(1,39)=11,183, p<0,002), тета-1- (F(1,39)=4,184, p<0,048), тета-2- (F(1,39)=11,577, p<0,002) и гамма- (F(1,39)=9,667, p<0,003) диапазонах. В тета-1- и гамма-диапазонах медитаторы и контрольные испытуемые обнаружили сходную топографическую и временную структуру реакции ВС на эмоциогенные стимулы: эффекты эмоциогенности стимула проявляются преимущественно в задних областях коры в период 0-600 мс. Однако, в дельта-диапазоне в теменно-височных-затылочных областях коры обнаружены специфические межгрупповые различия, связанные с эмоциогенностью стимула и функциональной полушарной асимметрией (F(12,468)=3,803, p<0,009). В ответ на положительные и отрицательные стимулы медитаторы демонстрируют меньшие значения ВС билатерально по всему временному интервалу. Наряду с этим, наблюдаемая у контрольных испытуемых более выраженная синхронизация в правом полушарии в ответ на негативные эмоциогенные стимулы в интервале 400-600 мс. от начала предъявления отсутствует у медитаторов. Таким образом, на субъективном уровне эмоциогенные сигналы оказывают меньшее активационное воздействие на медитаторов по сравнению с контрольными испытуемыми. Процесс эмоционального восприятия у медитаторов характеризуется более "экономным" использованием активационных ресурсов: вне зависимости от категории стимула, медитаторы демонстрируют более низкие значения синхронизации в дельта-, тета-1-, тета-2- и гамма-диапазонах. В дельта-диапазоне в теменно-височно-затылочных областях коры билатерально у медитаторов наблюдаются более низкие значения связанной с эмоциогенностью вызванной синхронизации. Кроме того, характерная для контрольных испытуемых более выраженная синхронизация в правом полушарии в ответ на негативные эмоциогенные стимулы, отсутствует у медитаторов. 4) Спектры мощности ЭЭГ в модели переживания дискретных эмоций. А) Субъективный отчёт. До начала стимуляции значимых различий в эмоциональном профиле контрольных испытуемых и медитаторов обнаружено не было. Внутригрупповые анализы подтвердили наличие роста значений "целевых" эмоций (например, радость при индукции радости) при предъявлении эмоциональных клипов по сравнению с нейтральными в обеих группах испытуемых. Однако, на межгрупповом уровне медитаторы демонстрировали значимо менее выраженную интенсивность переживания эмоций радости (F(9,486)=4,51, p<0,001), гнева (F(9,486)=6,28, p<0,001), отвращения (F(9,486)=2,70, p<0,005) и грусти (F(9,486)=3,80, p<0,001). Б) Спектры мощности ЭЭГ. Эмоция радости. Переживание эмоции радости вне зависимости от фактора группы сопровождалось ростом тета-1- (F(1,55)=10,760, p<0,002) и бета-2- (F(1,55)=5,610, p<0,021) мощности по всему корковому плану, а также ростом бета-3- (F(4,220)=4,400, p<0,002) и гамма- (F(2,110)=5,350, p<0,006) мощности в центральных и задних отделах коры. Кроме того, у испытуемых обеих групп в альфа-1-диапазоне наблюдалось снижение мощности в центральных и задних областях (F(4,220)=9,250, p<0,001). Анализ взаимодействия ГРхЭУхОК (F(2,110)=5,997, p<0,008) в тета-2-диапазоне указывает на рост мощности в задних областях коры у контрольных испытуемых и отсутствие значимых изменений у медитаторов. Напротив, в альфа-2-диапазоне взаимодействие ГРхЭУ (F(1,55)=8,520, p<0,005) обусловлено снижением мощности у медитаторов и отсутствием изменений мощности в группе контроля. В бета-1-диапазоне взаимодействие ГРхЭУ (F(1,55)=5,352, p<0,024), как и в случае тета-2-полосы, обеспечивается реактивностью контрольных испытуемых и ареактивностью медитаторов. В) Эмоция гнева. В обеих группах переживание эмоции гнева сопровождалось билатеральным усилением тета-2-мощности (F(2,110)=12,130, p<0,001) в центральных и задних областях коры. В бета-1-диапазоне наблюдался асимметричный рост мощности в левом полушарии (F(1,55)=5,897, p<0,018). Характерной особенностью медитаторов явилось отсутствие наблюдаемого у контрольных испытуемых роста бета-2- (F(2,110)=4,213, p<0,029), бета-3- (F(2,110)=6,814, p<0,002) и гамма- (F(2,110)=5,218, p<0,007) мощности в передних областях коры. Г) Эмоция отвращения. У обеих групп эмоция отвращения характеризовалась эффектами альфа-1- (F(2,110)=21,003, p<0,001) и альфа-2- (F(2,110)=8,550, p<0,001) десинхронизации в центральных и задних областях коры. В тета-2-диапазоне (F(1,55)=5,346, p<0,025) медитаторы не обнаруживают значимых изменений мощности, в то время как у контроля мощность растёт по всему корковому плану. В бета-1-диапазоне (F(2,110)=3,392, p<0,041) в задних областях коры у медитаторов наблюдается снижение мощности, а у контрольных - отсутствие реакции. Д) Эмоция грусти. Эмоция грусти характеризовалась у обеих групп испытуемых усилением альфа-2- (F(2,110)=4,327, p<0,026) и бета-1- (F(4,220)=9,331, p<0,001) мощности преимущественно в задних отведениях коры. В тета-2-диапазоне (F(1,55)=8,175, p<0,006) у медитаторов отсутствует наблюдаемый у контроля рост мощности. В отличии от контроля медитаторы демонстрируют снижение альфа-1-мощности (F(2,110)=4,163, p<0,018) в задних областях коры. В высокочастотной области спектра (бета-3- F(1,55)=4,414, p<0,041; гамма - F(2,110)=3,660, p<0,045) только контрольные испытуемые демонстрируют значимые изменения - десинхронизацию в центрально-теменных областях коры. Таким образом, на субъективном уровне медитаторы демонстрируют меньшую интенсивность переживания вызванных дискретных эмоций. По данным ЭЭГ, переживание вызванных положительных и отрицательных эмоций медитаторами по сравнению с контролем характеризуется: отсутствием эффектов усиления тета-2-мощности в задних (эмоция радости) либо во всех (эмоция грусти и отвращения) областях коры; снижением бета-1-мощности в задних (эмоция отвращения) и отсутствием её усиления (эмоция радости) в передних и задних корковых областях; отсутствием роста бета-3- и гамма-мощности (эмоция гнева) в передних областях коры головного мозга и снижения (эмоция грусти) - в центрально-теменных областях. 5) Спектры мощности ЭЭГ и вегетативная реактивность (ЭКП) в условиях стрессовой стимуляции. А) Субъективный отчёт. На субъективном уровне, в условиях стрессовой стимуляции у обеих групп испытуемых наблюдался значимый рост интенсивности переживаний отрицательных эмоций (гнева, страха, тревоги, отвращения, отторжения), однако, у медитаторов амплитуда реакций была достоверно ниже (F(9,477)=12,44, p<0,001) Б) Спектры мощности ЭЭГ. Стрессовая стимуляция в обеих группах приводила к альфа-1- (F(2,106)=10,748, p<0,001), альфа-2- (F(2,106)=6,188, p<0,004) десинхронизации в центрально-теменных и гамма- (F(2,106)=4,508, p<0,027) синхронизации в передне-центральных областях коры. В бета-1-диапазоне (F(1,53)=5,237, p<0,026) медитаторы обнаруживают выраженную десинхронизацию, в то время как у контроля значимые изменения в этой частотной полосе отсутствуют. В) Электрокожный потенциал. На вегетативном уровне стрессовая стимуляция приводила к достоверно более выраженному росту амплитуды уровня электрокожного потенциала у контрольных испытуемых по сравнению с медитаторами (p<0,01). А по данным внутригрупповых анализов, изменения электрокожного потенциала в ответ на стрессовый вызов по сравнению с условием нейтральной стимуляции были значимыми только в группе контроля (p<0,04). Помимо этого были обнаружены положительные корреляции между изменением спектральной бета-1-мощности ЭЭГ и амплитудой электрокожных реакций (диапазон от r=0,27 до r=0,42) в условиях стрессовой стимуляции. Таким образом, по данным субъективных оценок в условиях психоэмоционального стресса медитаторы характеризовались более низкой интенсивностью переживаний эмоций отрицательного круга. Стрессозащитные эффекты медитации на вегетативном уровне подтверждаются более низкими уровнями симпатической активации по данным электрокожной активности, а на корковом отражаются в снижении активности ЭЭГ в бета-1-диапазоне. Результаты корреляционного анализа подтверждают связь между изменениями мощности ЭЭГ в бета-диапазоне и амплитуды ЭКП: более выраженное снижение мощности ЭЭГ ассоциировалось с минимизацией реакций ЭКП на стрессовый вызов. ВЫВОДЫ По сравнению с контрольными испытуемыми на личностном уровне медитаторы (медитация по системе Сахаджа Йога), обладая большей интровертированностью, отличаются более высокой психоэмоциональной стабильностью и лучшими способностями к идентификации и выражению эмоций. В состоянии физиологического покоя по данным ЭЭГ медитаторы демонстрируют более высокие значения тета-1- и тета-2-мощности в передних, а альфа-1-мощности - во всех областях коры. В диапазоне гамма-активности наблюдается общее снижение мощности по всему корковому плану. По данным значений электрокожного потенциала и вариативности сердечного ритма медитаторы обнаруживают сниженные уровни вегетативной активации. Состояние медитации характеризуется усилением спектральной мощности в тета-1-, тета-2- и альфа-1-диапазонах наряду с фокусированным снижением оценок фрактальной размерности ЭЭГ в передне-медиальных областях коры, а также ростом длиннодистантной тета-когерентности между передней и задней ассоциативной корой с формированием "центра тяжести" в левой префронтальной области. Субъективные показатели положительных эмоциональных переживаний в состоянии медитации положительно коррелируют с изменением тета-1-мощности, а торможение когнитивной активности - с изменением тета-2-мощности в передне-медиальных областях коры. Процесс восприятия у медитаторов отличается более "экономным" использованием активационных ресурсов. Межгрупповые различия эмоционального восприятия обнаруживаются в диапазоне дельта-активности в теменно-височно-затылочных областях коры: в ответ на положительные и отрицательные эмоциональные стимулы по сравнению с нейтральными медитаторы демонстрируют меньшие значения вызванной синхронизации. Наряду с этим, у медитаторов отсутствует наблюдаемая у контрольных испытуемых более выраженная синхронизация в правом полушарии в ответ на негативные эмоциогенные стимулы. На субъективном уровне медитаторы демонстрируют меньшую интенсивность переживания вызванных дискретных эмоций. Нейрофизиологически, особенности реализации эмоций отражаются в изменениях реактивности в тета-2-, бета-1-, бета-3- и гамма-диапазонах. В условиях психоэмоциональной стрессовой стимуляции защитные эффекты медитации на вегетативном уровне подтверждаются более низкими уровнями симпатической активации по данным электрокожной активности, а на корковом - отражаются в снижении мощности ЭЭГ в бета-диапазоне. Список сокращений: ВД/ВС - вызванная десинхронизация/синхронизация; ВСР - вариативность сердечного ритма; ЭКГ - электрокардиограмма; ЭКП - электрокожный потенциал; ЭЭГ - электроэнцефалограмма; DCx - фрактальная размерность. |
